Av Hans Fredrik Sunde
Hans Fredrik er doktorgradskandidat på Senter for Fruktbarhet og Helse, som er en del av Folkehelseinstituttet. Her forsker han på sammenhengen mellom mental helse og sosiale forskjeller på tvers av generasjoner. Fra tidligere har Hans Fredrik en mastergrad i psykologi fra NTNU i Trondheim. Du kan lese denne og andre tekster i vår utgave om mangfold, som kommer ut snart!
Jo lenger tilbake i tid du går, jo større blir familien din. Hvor langt tilbake må du før du er i slekt med alle? Svaret er overraskende kort: Bindeleddet mellom deg og alle andre levende mennesker levde antagelig for 3600 år siden, lenge etter at for eksempel pyramidene i Giza ble bygget. Hvordan kan dette stemme, og hva slags konsekvenser har dette for hvordan vi tolker betydningen av genetiske forskjeller?
Våre intuisjoner er til begrenset hjelp når det kommer til slektskap. Vi må derfor starte med litt enkel matte. Alle har to foreldre. Dine foreldre har også to foreldre hver. Det betyr at du har fire besteforeldre. De igjen har to foreldre hver, som betyr at du har åtte oldeforeldre. Så langt så godt. Men så er spørsmålet: Hvor langt tilbake i tid må du før du har 10 000 forfedre? Hva med 100 000 forfedre? Eller en million forfedre?
Antall forfedre i hver generasjon øker eksponentielt, en matematisk finurlighet som ofte tar vår intuisjon på senga. For å finne ut hvor mange forfedre, \(X\), du har et gitt antall generasjoner tilbake, \(N\), kan du bruke en enkel eksponent: \(X = 2^N\) For eksempel, for å finne ut hvor mange forfedre du har fire generasjoner tilbake i tid – det vil si antall tipp-oldeforeldre – kan du regne ut \(2^4 = 16\). Du hadde kanskje ikke trengt en formel for å finne ut det, men det vil komme godt med.
En generasjonslengde er aldersforskjellen mellom foreldre og barn. I gjennomsnitt er det omtrent 30 år (Fenner, 2005; Tremblay & Vézina, 2000). Foreldrene dine var altså på din alder for omtrent 30 år siden, og besteforeldrene dine var på din alder 30 år før der igjen. Hvis vi fortsetter var dine oldeforeldre like gamle som deg for omtrent 90 år siden, og dine tipp-oldeforeldre for omtrent 120 år siden. Med andre ord tar det rundt 120 år for fire generasjoner å passere. Med både en generasjonslengde og en måte å telle antall forfedre på er vi nå klare for å besvare spørsmålet: Hvor langt tilbake i tid må du før du har 10 000 forfedre? Forsøk å gjette på et svar før du leser videre. Hva sier intuisjonen deg?
Vi har nå to muligheter. Vi kan prøve oss frem med flere tilfeldige antall generasjoner helt til vi finner svaret, noe som kan være spennende og terpende for intuisjonene våre. En snarvei er derimot å bruke logaritmer. Logaritmer er til eksponenter hva divisjon er til multiplikasjon: et speilbilde! Logaritmer lar deg finne eksponenten du må bruke for å komme til et gitt tall. Siden grunntallet vårt er 2 (to foreldre per person) blir formelen
\(N = \log_2(X)\), hvor \(X\) er antall forfedre, og \(N\) igjen refererer til antall generasjoner tilbake i tid.
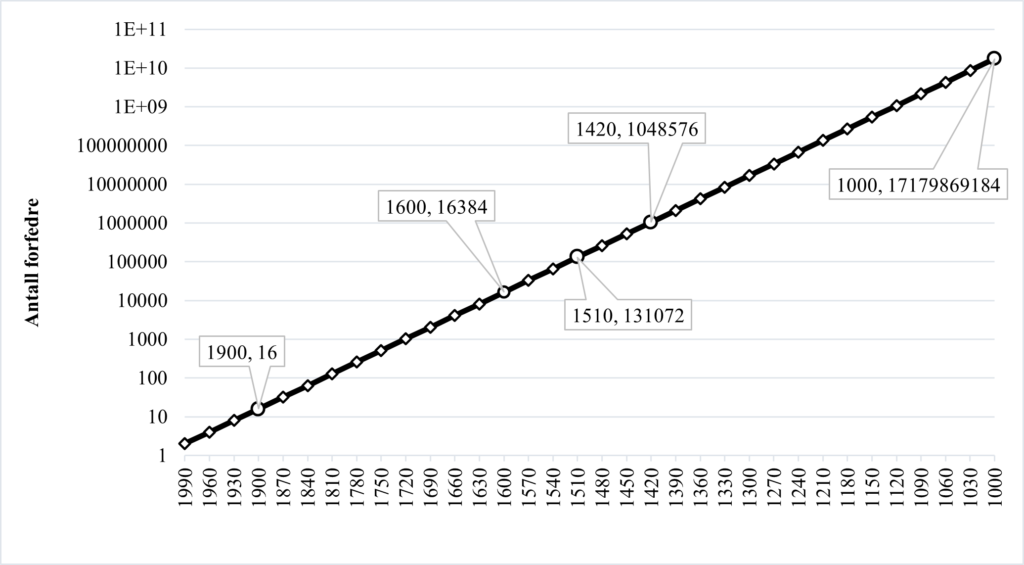
Hvor mange generasjoner må du tilbake i tid for å ha 10 000 forfedre? Svaret er \(\log_2(10 000) = 13.29\). Med andre ord passerer man 10 000 forfedre et sted mellom 13 og 14 generasjoner tilbake. Det vil si at du har over 10 000 tipp-tipp-tipp-tipp-tipp-tipp-tipp-tipp-tipp-tipp-tipp-oldeforeldre. Når var de på din alder? For å finne ut det ganger vi antall generasjoner med 30 år, og finner at de var på din alder for omtrent 400 år siden. Du hadde 16 forfedre på din alder for 120 år siden, men kun 300 år før det hadde du over 10 000!
Hvor langt må du tilbake før du har 100 000 forfedre? Da må du mellom 16 og 17 generasjoner tilbake i tid, altså rundt 500 år. For å gå fra 10 000 til 100 000 forfedre må du altså bare 100 år lenger tilbake i tid, til starten av 1500-tallet. Til sammenligning var det omtrent 170 000 innbyggere i Norge på den tiden (Sandmo, 2015). Du ser mønstret nå: Antall forfedre per generasjon øker fort. Veldig fort. For å komme til en million forfedre må du ikke lenger tilbake enn 1400-tallet. Da slaget stod på Stiklestad for 1000 år siden hadde du omtrent ti milliarder forfedre.
Det er selvfølgelig ikke snakk om ti milliarder unike personer. Vi snakker om ti milliarder unike roller. Du skal ikke mange generasjoner tilbake før du har personer som innehar flere roller, for eksempel ved at de finnes på både mors og fars side. For de 8.5 prosentene av jordens befolkning som er barn av søskenbarn eller tremenninger vil enkelte oldeforeldre, og noen ganger også besteforeldre, være på begge sider av familietreet (Modell & Darr, 2002). Andre må noe lenger tilbake, men til syvende og sist er alle et resultat av innavl. Det er ikke flere mennesker å ta av. Grenene kveiler seg tilbake inn i familietreet og danner et nett av slektskap.
Antagelig har familietreet kveilet seg inn i seg selv flere ganger før det når størrelsen på en befolkning. Dette er fordi det ikke er tilfeldig hvem folk får barn med. Folk parer seg som regel innenfor geografiske skiller, som begrenser mulige personer å få barn med. I teorien er det mulig for en populasjon å være genetisk isolert ved at ingen får barn med noen utenifra, men det er sjelden slik i praksis. Det skal ikke mange eventyrlystne og amorøse personer til før den genetiske boblen sprekker. Fremmede på besøk i området her, en handelsreise der, også videre. Dette lar gener spre seg mellom populasjoner, og gjør at du finner slektskapsstier som etter hvert knytter sammen hele jordens befolkning.
Det er ingen nyhet at vi alle er i slekt med hverandre. Går vi langt nok tilbake er vi i slekt med aper, amfibier, og amøber. 10 millioner år må du tilbake før du finner bindeleddet mellom mennesket og sjimpanser (Arnason et al., 1998), men hvor langt tilbake må du for å finne bindeleddet mellom deg og et tilfeldig annet menneske?
Forsøk på å besvare dette spørsmålet har ofte kun sett på slektstre som utelukkende klatrer opp på enten mors eller fars side. Dette gjøres ved genetiske analyser av mitokondria, som alltid arves fra mor, og Y-kromosomet, som alltid arves fra far. Søket etter Mitochondrial Eve og Y-chromosomal Adam tyder på at hun levde for mellom 99 000 – 148 000 år siden og han mellom 120 000 – 156 000 år siden (Poznik et al., 2013). Hvor langt må du tilbake hvis du kan bruke begge sider av familietreet samtidig?
Både matematiske modeller og genetiske undersøkelser tyder på at du finner en som er forfar (eller mor!) til nesten alle nålevende europeere for mindre enn 1000 år siden (Chang, 1999; Ralph & Coop, 2013). Vi mangler enda gode nok genetiske data for å gjøre tilsvarende analyser for hele verdens befolkning, men de matematiske modellene tyder på at det yngste bindeleddet mellom deg og alle levende mennesker – la oss kalle denne personen Primus – levde for omtrent 3600 år siden (Rohde et al., 2004). Det er 1000 år etter pyramidene i Giza ble bygget.
Det høres kanskje usannsynlig kort ut, men det skal ikke mye seksuell kontakt til mellom selv nokså isolerte populasjoner før gener raskt sprer seg i de påfølgende generasjonene. Regnestykket for antall etterkommere er tross alt det samme som antall forfedre, selv om det kompliseres av variasjon i antall barn. Menneskearten krysset Beringstredet over til Amerika for første gang over 15 000 år siden, men det betyr ikke at kontinentet har vært genetisk isolert siden. Den matematiske modellen nevnt over antar kun at én persons gener krysset Beringstredet hver 10. generasjon (Rohde et al., 2004). Det er nok til å knytte slektskapsstier nedover kontinentet. Uansett er det nok få individer i dagens Latin-Amerika som ikke har minst en europeisk forfar fra Colombus sin tid. Europeernes inntog på kontinentet for 500 år siden har satt tydelige genetiske spor (Chacón-Duque et al., 2018).
Alle levende mennesker er etterkommer av Primus, men Primus er ingen Adam eller Eva. Primus levde samtidig med mange andre mennesker som også har etterkommere i dag. Det spesielle med Primus er at hans eller hennes millioner av etterkommere har paret seg på kryss og tvers, slik at du kan tegne en sti fra Primus til alle som lever i dag. Selv om denne rollen tilhører en spesifikk person, er den ikke dermed låst. For eksempel kan en senere person innta rollen hvis en av slektsgrenene dør ut.
Primus sine forfedre vil også være forfedre for hele jordens befolkning, og jo lenger bakover du går, desto flere er det som har Primus som etterkommer. Etter hvert kommer du til et punkt hvor alle dalevende personer med levende etterkommere i dag er forfedre til alle som lever i dag. Fra dette punktet og bakover vil alle nålevende personer ha et identisk sett med forfedre. Matematiske modeller anslår at det er rundt år 1000 siden hvis du begrenser deg til Europeere, og mellom 5000 og 15 000 år siden for hele jordens befolkning (Ralph & Coop, 2013; Rohde, 2003; Rohde et al., 2004).
Disse kuriositetene er interessante i seg selv, men det er en del viktige konsekvenser som følger. En av de er betydningen av genetiske forskjeller. Fra mental helse og intelligens, til personlighet og holdninger: Grunnen til at du ikke er lik naboen din skyldes i stor grad genetiske forskjeller (Harden, 2020; Polderman et al., 2015; Turkheimer, 2000). Rene miljøfaktorer er selvfølgelig også viktige, men de er som regel enten små eller usystematiske. Genetiske forskjeller er den største systematiske faktoren som gjør oss forskjellig fra hverandre.
Siden vi har arvet de fleste av våre genvarianter fra våre forfedre kan det være fristende å konkludere at det samme gjelder forskjeller mellom populasjoner og ætter, men dette er feil. Utover din nærmeste familie, er ikke din spesifikke genetiske historie – din slektstavle – så viktig for å forstå hvorfor du er forskjellig fra andre. Selv blant helsøsken, som i snitt deler 50% av sitt unike DNA, er det de genetiske forskjellene som er den viktigste forklaringsfaktoren. Genetisk slektskap vannes fort ut for hver generasjon tilbake. Kun 12.5% av ditt unike DNA kan spores til en spesifikk oldemor, kun 6% til en tipp-oldemor, og kun 3% til en tipp-tipp-oldemor. Den lille genetiske likheten som er igjen drukner fort i en sjø av genetisk ulikhet, og man vil fort finne forfedre som genetisk sett like gjerne kunne vært fremmede. Menneskeartens paringsmønstre fungerer som en kjøkkenmaskin, og vil effektivt blande gener over relativt korte tidsperioder. Det er ikke arven etter slekten din, men snarere din unike kombinasjon av gener som gjør deg til den du er.
I tillegg til at genetisk likhet vannes ut når man klatrer familietreet, har vi sett at familietreet sprer seg ut fort, og at ingen populasjoner lever i en genetisk boble. Paringer på tvers av landegrenser og kulturer åpner slektskapsstier på kryss og tvers, og fører til at nye genetiske varianter ikke holdes isolerte i enkelte populasjoner. Den genetiske kjøkkenmaskinen gjør at genetisk variasjon mellom populasjoner også vannes ut. Innad i populasjoner derimot vil den genetiske variasjonen opprettholdes av stadig nye og unike genetiske mutasjoner. Derfor finner vi mer genetisk variasjon innad i populasjoner enn mellom populasjoner (Witherspoon et al., 2007). Genetiske forskjeller er altså viktige for å forstå forskjeller mellom individer innad i en populasjon, men ikke forskjeller mellom populasjoner.
Genetikk har dermed både en samlende side og en skillende side: Det knytter alle i hele verden sammen gjennom et nettverk av slektskap og et relativt nylig bindeledd, men det gjør deg også til en unik person, annerledes enn alle de andre. Genetikk viser oss både hvor like og hvor forskjellige vi er.
PS: Det er andre interessante og viktige konsekvenser knyttet til matematikken bak slektskap og forfedre, men det er hverken tiden eller plass til alt her. Isteden må jeg henvise deg til andre kilder, for eksempel inspirasjonen bak det meste av denne teksten, Adam Rutherfords A Brief History of Everyone Who Ever Lived (2016).
Referanser
Arnason, U., Gullberg, A., & Janke, A. (1998). Molecular Timing of Primate Divergences as Estimated by Two Nonprimate Calibration Points. Journal of Molecular Evolution, 47(6), 718-727. https://doi.org/10.1007/pl00006431
Chacón-Duque, J.-C., Adhikari, K., Fuentes-Guajardo, M., Mendoza-Revilla, J., Acuña-Alonzo, V., Barquera, R., Quinto-Sánchez, M., Gómez-Valdés, J., Everardo Martínez, P., Villamil-Ramírez, H., Hünemeier, T., Ramallo, V., Silva De Cerqueira, C. C., Hurtado, M., Villegas, V., Granja, V., Villena, M., Vásquez, R., Llop, E., … Ruiz-Linares, A. (2018). Latin Americans show wide-spread Converso ancestry and imprint of local Native ancestry on physical appearance. Nature Communications, 9(1). https://doi.org/10.1038/s41467-018-07748-z
Chang, J. T. (1999). Recent Common Ancestors of All Present-Day Individuals. Advances in Applied Probability, 31(4), 1002-1026. http://www.jstor.org/stable/1428340
Fenner, J. N. (2005). Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies. American Journal of Physical Anthropology, 128(2), 415-423. https://doi.org/10.1002/ajpa.20188
Harden, K. P. (2020). “Reports of My Death Were Greatly Exaggerated”: Behavior Genetics in the Postgenomic Era. Annual Review of Psychology. https://doi.org/10.1146/annurev-psych-052220-103822
Modell, B., & Darr, A. (2002). Genetic counselling and customary consanguineous marriage. Nature Reviews Genetics, 3(3), 225-229. https://doi.org/10.1038/nrg754
Polderman, T. J. C., Benyamin, B., De Leeuw, C. A., Sullivan, P. F., Bochoven, A. V., Visscher, P. M., & Posthuma, D. (2015). Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nature Genetics, 47(7), 702-709. https://doi.org/10.1038/ng.3285
Poznik, G. D., Henn, B. M., Yee, M.-C., Sliwerska, E., Euskirchen, G. M., Lin, A. A., Snyder, M., Quintana-Murci, L., Kidd, J. M., Underhill, P. A., & Bustamante, C. D. (2013). Sequencing Y Chromosomes Resolves Discrepancy in Time to Common Ancestor of Males Versus Females. Science, 341(6145), 562-565. https://doi.org/10.1126/science.1237619
Ralph, P., & Coop, G. (2013). The Geography of Recent Genetic Ancestry across Europe. PLoS Biology, 11(5), e1001555. https://doi.org/10.1371/journal.pbio.1001555
Rohde, D. L. T. (2003). On the Common Ancestors of All Living Humans. http://faculty.collin.edu/dkatz/Rohde-MRCA-two.pdf
Rohde, D. L. T., Olson, S., & Chang, J. T. (2004). Modelling the recent common ancestry of all living humans. Nature, 431(7008), 562-566. https://doi.org/10.1038/nature02842
Rutherford, A. (2016). A Brief History of Everyone Eho Ever Lived: The Stories in Our Genes. Weidenfeld & Nicolson.
Sandmo, E. (2015, 02. nov 2020). Livet og Pesten. https://www.norgeshistorie.no/kirkestat/1112-livet-og-pesten.html
Tremblay, M., & Vézina, H. (2000). New Estimates of Intergenerational Time Intervals for the Calculation of Age and Origins of Mutations. The American Journal of Human Genetics, 66(2), 651-658. https://doi.org/10.1086/302770
Turkheimer, E. (2000). Three Laws of Behavior Genetics and What They Mean. Current Directions in Psychological Science, 9(5), 160-164. https://doi.org/10.1111/1467-8721.00084
Witherspoon, D. J., Wooding, S., Rogers, A. R., Marchani, E. E., Watkins, W. S., Batzer, M. A., & Jorde, L. B. (2007). Genetic Similarities Within and Between Human Populations. Genetics, 176(1), 351-359. https://doi.org/10.1534/genetics.106.067355
